|
Племенное разведение собак
|
|
| Svarta | Дата: Четверг, 26.04.2012, 18:56 | Сообщение # 91 |
 Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
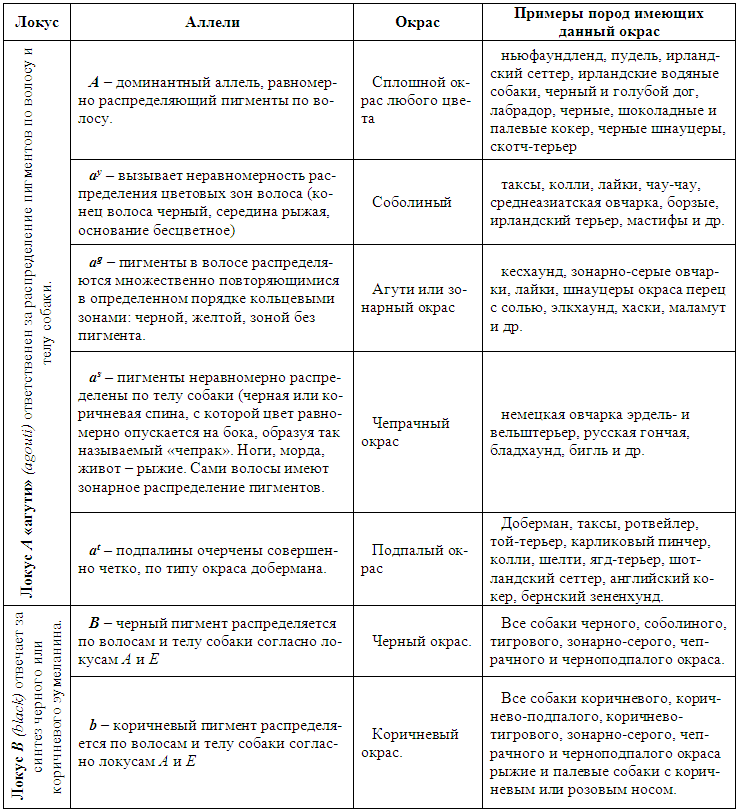
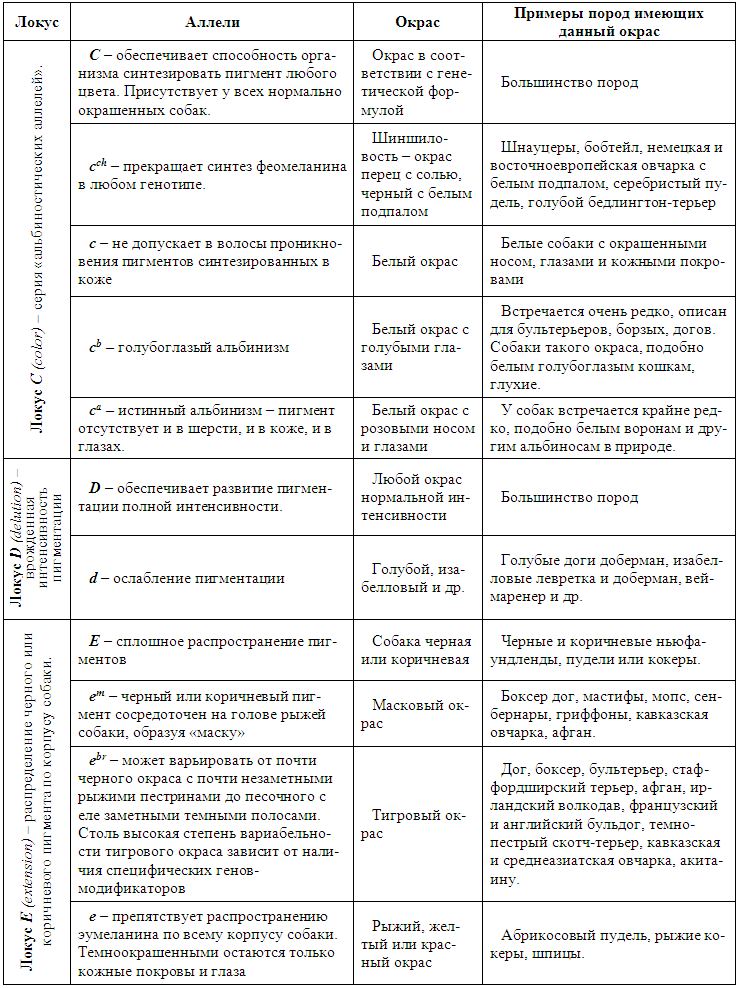
| Качественные и количественные признаки.
Все признаки, которыми обладают живые организмы, принято делить на две категории — качественные и количественные. Качественные — признаки имеющие четко различимые формы, например, окраска или генетические аномалии, передаваемые отдельными генами. На фенотипическое проявление качественных признаков условия среды практически не влияют. Для характеристики популяции по качественным признакам используют понятия частота генов и генотипов.
Однако большая часть свойств организма представлена количественными признаками . Они проявляют в основном непрерывную изменчивость и могут быть измерены — рост, длина шерсти, вес. Количественные признаки, в большей степени, чем качественные, зависят от условий среды и обусловлены многими генами, так называемыми полигенами, то есть системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называется полимерным. Эти гены также называются аддитивными, так как их действие суммируется.
Распределение их численных значений в популяции приближается к кривым нормального распределения. Их наследование можно рассматривать по схеме полигибридного скрещивания.
Селекционеру приходится иметь дело в основном с непрерывной изменчивостью. Менделевский подход к изучению количественных признаков затруднен, хотя они и подчиняются тем же законам классической генетики, что и качественные.
Взаимодействие неаллельных генов
Гены, находящиеся в разных локусах также могут влиять друг на друга. При этом можно выделить несколько типов подобного взаимодействия.
Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект действия других генов, называются генами-модификаторами. Изучение окраски у млекопитающих показало, что наряду с крайними формами, обладающими полным развитием пигмента или его отсутствием, наблюдается целый ряд генотипически обусловленных форм. Так, белая пятнистость у собак варьирует от нескольких белых волосков на месте первичной точки депигментации до полностью белой собаки с небольшим пучком окрашенных волос в одном из пигментных центров. В пределах генотипа, определяемого локусом белой пятнистости S, можно выделить массу переходных форм.

Рис. 17. Разные варианты белой пятнистости собак
Широкую вариабельность, обусловленную генами-модификаторами, имеют чепрачный и тигровый окрасы собак.
Если признак образуется при наличии двух пар неаллельных генов, дающих при совместном действии не тот эффект, который каждый из них обусловливает самостоятельно, то такие аллели обозначают как комплементарные (дополняющие друг друга). В качестве примера комплементарности у собак обычно приводят взаимодействие генов локусов В и Е, определяющих окраску.

Рис. 18. Комплементарное взаимодействие генов, определяющих окраску собак: соотношение — 9 черных (Ч): 3 коричневых (К): 3 рыжих (Р): 1 палевая (П)
Гены локуса В отвечают за синтез черного ( В ) или коричневого ( b ) пигмента. Гены локуса Е отвечают за распределение этих пигментов. Аллель Е способствует распространению черного или коричневого пигмента по всему телу собаки. Аллель е препятствуют их синтезу в шерстном покрове. Собаки с генотипом ее — рыжие или желтые. Черный или коричневый пигмент при этом сосредоточен только на кожных покровах морды собаки.
Формирование окраски собаки зависит от присутствия обоих пар генов. Собаки с генотипом ЕЕ или Ее — черные или коричневые в зависимости от аллелей B или b. При Е-ВВ или Е-Bb — собака черная, при E-bb — коричневая.
Собаки с генотипом ееВ- — рыжие с черным носом. Собаки генотипа bbee — обычно палевые или светло-желтые со светлым носом.
Комплементарным взаимодействием генов обусловлен особый тип паралича задних конечностей у помесей датского дога с сенбернаром. Генетический анализ проведенный Стокардом (Stockard, 1936), показал, что при чистопородном разведении как датских догов, так и сенбернаров паралич не развивается.
Аналогичное заболевание отмечалось у некоторых помесных собак-ищеек (Петухов и др., 1985).
В любой паре аллелей доминантный ген препятствует (полностью или частично) проявлению своего рецессивного партнера. Но иногда действие доминантного аллеля подавляется действием гена из другого локуса. Всемогущий ген, который блокирует действие другого гена или генов называется эпистатичным. А само явление — эпистазом. Гены, действие которых подавлено, называется гипостатичными .
Так, рецессивные гены окраски собак из локуса C не позволяют синтезироваться пигментам, определяющим окраску шерсти. Собака гомозиготная по ним — белая.
Один и тот же мутантный признак может проявляться у одних и не проявляться у других особей родственной группы. Способность данного гена проявлять себя фенотипически называется пенетрантностью. Пенетрантность определяется по проценту особей в популяции, имеющих мутантный фенотип. При полной пенетрантности (100%) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100%) ген проявляется фенотипически не у всех особей.
У собак достаточно часто встречаются видоизменения хвостов в виде их укороченности, разнообразных изломов и изгибов. Можно предположить, что многообразие этого признака обусловлено его неполной пенетрантностью.
Степень пенетрантности может сильно изменяться под воздействием условий среды.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 18:57 | Сообщение # 92 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| 
Рис. 19. Схема дигибридного расщепления с рецессивным эпистазом: вВ F 2 получено 9 черных: 3 коричневых: 4 белых собаки. Таким образом, наблюдается характерное для рецессивного эпистаза отклонение от теоретически ожидаемого расщепления 9:3:3:1
Часто особи, обладающие тем же генотипом в отношении какого-либо наследственного признака, очень сильно различаются по его экспрессивности, то есть степени проявления данного признака. Один и тот же ген у разных особей в зависимости от влияния генов-модификаторов и внешней среды может проявить себя фенотипически по-разному. Внешняя среда и гены-модификаторы могут изменить экспрессию гена, то есть выражение признака.
В отличие от пенетрантности, которая указывает, у какой доли особей в популяции проявляется данный признак, экспрессивность относится к изменчивости признака у тех особей, у которых он проявляется. Так у собак экспрессивность развития прибылых пальцев варьирует от полностью развитых пальцев на обеих задних конечностях до наличия их в зачаточном состоянии только на одной конечности. Подобная вариация экспрессивности характерна и для других наследуемых признаков, в частности и для вышеупомянутых хвостов.
Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Достаточно широко распространено явление плейотропии — влияния одного гена на развитие двух и более признаков. Классический «собачий» пример плейотропного влияния — действие фактора Мерля, (локус М; окраса собак). Аллель M в гетерозиготном состоянии Мm дает пятнистость типа «арлекин» характерную для догов. Аллель M в гетерозиготе Mm в сочетании с подпалом дает «мраморный» (blue-merle) окрас, типичный для колли и шелти. В гомозиготном состоянии ММ он ведет к рождению чисто белых щенков ( white-merle ) со значительными аномалиями органов чувств. Такие щенки часто погибают еще до рождения, а если и рождаются живыми, то их жизнеспособность резко снижена.
Явление плейотропии объясняется тем, что гены плейотропного действия контролируют синтез ферментов, участвующих в многочисленных обменных процессах в клетке и в организме в целом и, тем самым, одновременно влияющих на проявление и развитие многих признаков.
Некоторые гены вызывают столь сильные отклонения от нормы, что снижают жизнеспособность организма или даже приводят его к гибели. Такие гены называют летальными, то есть смертоносными, или сублетальными — снижающими жизнеспособность. В большинстве случаев летальные гены полностью рецессивны, поэтому гетерозиготные носители этих генов фенотипически совершенно неотличимы от нормальных особей. В гомозиготном состоянии такие гены могут нарушать нормальный ход развития эмбриона на любых стадиях. О возможности наличия летальных генов можно косвенно судить по снижению средней численности пометов или по выпадению некоторой ожидаемой части фенотипов в расщеплении.
Так в случае скрещивания упоминавшихся выше черно-мраморных собак, гетерозиготных по фактору Мерля получается вместо ожидаемого соотношения 3:1, получается 2:1 т. е. 2 мраморных и 1 черная собаки Mm × Mm = ММ: 2 Mm: mm, где ММ белая нежизнеспособная собака. Белые щенки чаще не рождаются вовсе, так как погибают задолго до рождения.

Рис. 20. Наследование мраморности типа «арлекин» у догов. Ген М h (фактор Мерля) — доминантный с рецессивным летальным действием: 1 — скрещивание мраморных догов между собой; 2 — анализирующее скрещивание
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 18:58 | Сообщение # 93 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Фактор Мерля относится к категории доминантных летальных генов, которых значительно меньше, чем рецессивных. При желании его носителей легко изъять из разведения, так как они имеют характерный фенотип. Одни летальные гены обуславливают крупные аномалии, другие — нарушения физиологических процессов. Пути губительного воздействия большинства летальных генов не ясны. Таких генов может существовать сколь угодно много. Показано, что каждый человек является носителем в среднем 4–9 «вредных» или летальных генов. Аналогичных результатов можно ждать и по собакам. Известны летальные гены, которые при проявлении в эмбриональном состоянии, опасны и для жизни беременной суки, например, при наследственной контрактуре мышц плода, вследствие чего сука не может разродиться.
Взаимодействие генов, когда при их сочетании в одном организме развивается совершенно новая форма признака называется новообразованием.
Иногда новообразования приводят к появлению признаков дикого фенотипа. В этом случае их называют атавизмами, то есть возвратом к предковой форме или возвратом в дикий тип.
Частичные возвраты в дикий тип возможны при скрещивании двух особей одной породы, если эти производители происходят из удаленных друг от друга неродственных популяций. По-видимому, подобным образом можно объяснить и большое сходство между дворнягами, обитающими в самых разных местах.
Признаки, сцепленные с полом
Признаками, сцепленными с полом называют такие, которые формируются под воздействием генов, локализованных в Х -хромосоме. Наиболее характерным примером наследования признака сцепленного с полом является наследование гемофилии собак. У собак больных гемофилией, в крови не хватает фактора, который, взаимодействуя с кровяными пластинками (тромбоцитами), ускоряет превращение протромбина в тромбин. Гемофилия собак аналогична гемофилии человека и также обусловлена сцепленным с полом рецессивным геном. Ген, определяющий развитие гемофилии находится в Х -хромосоме и является рецессивным по отношению в нормальному аллелю. Следовательно, гемофилия проявляется только у гомозиготных самок (несущих этот ген в обеих Х -хромосомах) и гемизиготных самцов, несущих ген гемофилии в Х -хромосоме. Щенки-гемофилики обычно погибают в раннем возрасте от наружных или внутренних кровотечений. Сохранить до половозрелого состояния такого самца возможно только при постоянном введении специфических препаратов. Самки же неминуемо гибнут не позже первой течки. Гетерозиготные самки внешне абсолютно нормальны и плодовиты. Однако половина их детенышей мужского пола страдает гемофилией и половина детенышей-самок оказывается гетерозиготными по этому гену.
Родители:
сука-носительница гена гемофилии
ХHХh
кобель нормальный
XHY
— половая хромосома, несущая нормальный аллель

XH
Xh — половая хромосома, несущая ген гемофилии
К признакам сцепленным с полом относятся также врожденный гипотрихоз, отмеченный у такс и карликовых пуделей; мышечная дистрофия у ретриверов; синдром пошатывания, связанный с гипомиелинизацией и обнаруженный у чау-чау и ряда других пород; подвывих запястья; а также диафрагмальная грыжа, описанная у золотистых ретриверов.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 18:59 | Сообщение # 94 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Признаки ограниченные полом
Некоторые признаки совершенно независимо от места локализации генов их вызывающих, проявляются только у особей одного пола. Это так называемые ограниченные полом признаки. Таковы, например, дефекты развития половой системы, молочность и т. д. Одно из таких явлений — крипторхизм — непрохождение одного или обоих семенников через паховый канал в мошонку. Крипторхизм бывает двусторонний, право- или левосторонний и может быть вызван разными причинами: узостью пахового канала, короткими связками семенников, недоразвитием семенников. Неопустившиеся семенники могут находиться в разных местах брюшной полости. Крипторхизм может быть как врожденным, так и приобретенным. Среди разных его форм существует и генетически обусловленная. Однако из-за широкой вариабельности этого признака делать однозначные выводы о его природе невозможно. И уж совершенно неверно трактовать его как моногенный признак, локализованный в Х-хромосоме.
Закон гомологических рядов Н.И. Вавилова
Закон был сформулирован Н.И. Вавиловым в 1920 году. Н.И. Вавилов обнаружил, что все виды и роды, генетически близкие между собой характеризуются тождественными рядами наследственной изменчивости.
В основе закона гомологических рядов лежит параллелизм генотипической изменчивости у особей со сходным набором генов.
Этот закон имеет универсальный характер. Обнаружено сходство мутаций у разных видов животных. Так были отмечены проявления аналогичных форм аномалий у собак, кошек, кроликов, свиней, человека и др., что указывает на сходство строения многих ферментов и белков и соответственно на сходство генотипов. Таким образом, зная формы наследственных изменений у одного вида животных, можно предполагать, что они имеются или могут возникнуть и у другого близкого вида. Особенно тщательно изучены наследственные аномалии сельскохозяйственных животных и человека. У собак описано значительно меньше аномалий, однако это говорит лишь о меньшей изученности этого вида. Таким образом, при обнаружении новой аномалии у собак следует поинтересоваться, не описана ли она для других видов животных.
В основных породообразующих группах собак наблюдается гомология по многим признакам. Так, например, длина ног — ахондроплазия встречается у овчарок (вельшкорги), терьеров (скай терьеров, селихам терьеров, денди даймонт терьеров), гончих (бассет) легавых, догообразных (бульдоги), собак Тибета (лхасское апсо, ши тсу), периодически встречаются элементы ахондроплазии у пуделей. Подобных собак не отмечено только в группе борзых, поскольку этот признак является противоположным по отношению к акромегалии.
Во всех породных группах встречаются как гигантские, так и карликовые формы. У овчарок (комондор — шипперке), догообразных (мастиф — французский бульдог), терьеров (эрдельтерьер — той терьер), гончих (бладхаунд — бигль), шпицеобразных (аляска маламут — померанский шпиц), борзых (ирландский волкодав — левретка). Гомологическая изменчивость в размерах от гигантов до карликов наблюдается и среди узких породных групп. Например, шнауцеров (ризен — миттель — цверг), такс (стандартная — карликовая — кроличья), пуделей (стандартный — малый — карликовый — той пудель).
Признаками, проявляющими гомологию, являются окрас и тип шерсти.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:01 | Сообщение # 95 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Частная генетика собаки
Генетика окраски
Исследованием окраски собак занимались многие ученые. Сведения по генетике этого признака были опубликованы в монографиях Ильина (1932), Доусона (1937), Уитни (1947), Бернса и Фрезер (1966) и других. Многие авторы детально исследовали генетику окрасов собак одной породы.
В нашей стране экспериментальным изучением занимался в 30-е годы Н.А. Ильин в кинологической лаборатории и в Московском зоопарке.
Наиболее емким вкладом в проблему являются исследования К. Литтла, в течение многих лет изучавшего окрасы собак в Джексоновской лаборатории в США. Его работы шли в трех направлениях: экспериментальные скрещивания в лаборатории, анализ данных, полученных от заводчиков и владельцев питомников, племенная работа в своем питомнике. В течение 25 лет им было исследовано много тысяч пометов собак. Результатом явилась публикация монографии «The Inheritance of Coat Color in Dogs.» (1957).
Подробную сводку по окрасам собак приводит Р. Робинсон в монографии «Genetics for Dog Breaders» (1982).
Детальный анализ наследования окрасов собак дан в книгах М. Сотской «Пудель» (1998, 2004), Н. Московкиной и М. Сотской «Генетика и наследственные болезни собак и кошек» (2000).
Нужно отметить, что в работах разных авторов встречается несколько различающаяся символика в обозначении отдельных аллелей, а так же по-разному трактуются типы формирования того или иного окраса или места расположения отдельных аллелей в множественных сериях. Однако, в целом, разница во взглядах отдельных исследователей не носит принципиального характера.
Окрас шерстного покрова зависит от типа пигмента, формы пигментных гранул и распределения их по волосу. Все многообразие окрасок млекопитающих обусловлено наличием или отсутствием пигментов — меланинов .
Согласно современным представлениям меланин представлен двумя формами: эумеланином и феомеланином, которые существуют в виде пигментных зерен разнообразной формы. Восприятие цвета зависит от преломления света, проходящего через них или отражающегося от них. Эумеланин имеет две модификации: черный и коричневый. Феомеланин представлен желтым и красным вариантами.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:01 | Сообщение # 96 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| 

Таблица 9. Окраска собак
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:04 | Сообщение # 97 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Другие особенности экстерьера
Особенности экстерьера в основном являются полигенными и наследуются по типу промежуточного наследования, или как количественные признаки. При этом может проявляться как общая рецессивность, так и неполное доминирование. Многие из этих признаков проявляют неполную пенетрантность и экспрессивность. Некоторые сцеплены друг с другом. Поэтому менделевский анализ наследования морфологических особенностей в большинстве случаев оказывается затрудненным.
Строение тела собаки
Важные породообразующие признаки, связанные с формой головы, теле и длиной конечностей, обусловлены особенностями синтеза в организме собаки гормона роста — соматотропина (СТГ) и чувствительности к нему тканей организма. Среди множества пород собак можно выделить три группы: акромегалики , акромикрики и хондриодистрофики . Каждая имеет определенные фенотипические проявления динамического действия СТГ.
Акромегалики характеризуются вытянутыми в длину костями черепа и, частично, конечностей. Представителями этого типа являются различные борзые, колли и доберманы. Наследуется акромегалия по доминантному типу.
Акромикрики, наоборот, имеют укороченные кости лицевого черепа, так называемую мопсообразность, и, иногда, укороченные конечности. Эта группа представлена разными бульдогами, боксерами, пекинесами, мопсами, гриффонами и другими мопсообразными собаками. Выяснено, что акромикрия наследуется моногенно по рецессивному типу. Поэтому межпородные вязки приводят к потере типичного для этих пород облика.
При хондриодистрофии или иначе ахондроплазии задерживается рост трубчатых костей конечностей и развивается коротконогость, например, у таксы и вельшкорги. Подобные коротконогие собаки существовали еще в Древнем Египте и их изображения встречаются на древнеегипетских фресках. Хондриодистрофия также как акромикрия наследуется моногенно по рецессивному типу.
Коротконогость, возникающая за счет укорочения предплечий, по-видимому, имеет полигенную основу и может наследоваться по типу неполного доминирования. Аналогичным образом наследуется и укороченные бедро и голень, нарушающие правильный постав задних конечностей. Можно предполагать, что эти признаки имеют неполную пенетрантность и экспрессивность.
Исследования наиболее характерных для собак недостатков экстерьера показали, что многие признаки имеют отчетливо выраженный наследственный характер, однако могут усугубляться плохим выращиванием.
По-видимому, по доминантному типу наследуются такие признаки, как укороченная спина и связанная с ней куполообразная поясница, укороченный и скошенный круп (короче говоря, нарушение соотношения 2:1:1). Показано, например, что узкая грудная клетка доминирует над нормальной, коротконогость, возможно не полностью, над длинноногостью.
Отчетливо наследственный характер имеют углы сочленений.
Рост и вес тела собак представляют типичный пример количественных признаков. Как количественный признак наследуется и размер черепа.
Относительно же формы головы, как показывает практика, длинномордость доминирует над короткомордостью, широкий череп и нижняя челюсть над узкими, короткая теменная часть — над длинной. Метисы собак акромегалических пород (колли, борзые) имеют столь же длинную лицевую часть черепа, однако более широкую мозговую часть. Данный эффект сохраняется и в последующих поколениях.
Доминантным признаком с неполной пенетрантностью и экспрессивностью являются часто встречающиеся у собак прибылые пальцы на задних ногах.
Врожденное укорочение и дефекты хвоста могут быть обусловлены как доминантными, так и рецессивными генами, что подтверждается многочисленными наблюдениями. Гибриды бульдогов с собаками, имеющими хвосты нормальной формы, обычно обладают хвостами нормальной длины и формы.
Некоторые авторы указывают, что хвост саблевидной формы у собак доминирует над крючкообразным, кольцевидным и хвостом-поленом.
Часто, при скрещиваниях собак разных пород наблюдается появление признаков характерных для дикого типа, так называемый, уход в дикий тип. Это может быть объяснено неаллельностью генов, определяющих фенотип данных животных, а также наличием специфических регуляторных элементов генома. Частичные возвраты в дикий тип возможны и при скрещивании животных одной породы, происходящих из географически разобщенных популяций.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:05 | Сообщение # 98 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Зубная система
Строение зубной системы является исключительно важным элементом экстерьера собак. Значительные отклонения в форме, количестве зубов и прикусе может свидетельствовать о нарушениях генетической конституции не только отдельного животного, но и всей линии или семейства, к которому оно относится. Аномалии зубной системы наряду с другими аномалиями, например крипторхизмом, могут рассматриваться как маркеры гибридного дисгенеза.
История собаководства и многочисленные исследования селекционеров показывают, что в значительной мере наследственно обусловлена форма прикуса. Наследственно обусловлены врожденное укорочение нижней челюсти или прогнатия и укорочение верхней челюсти — прогения, также приводящие к образованию неправильного прикуса.
Большое внимание селекционеров привлекает также проблема полнозубости. Для некоторых пород наличие полного комплекта зубов является абсолютным. Для других — требования несколько смягчены. Врожденные аномалии количества зубов такие как олигодонтия и полиодонтия могут быть диагностированы как в момент актирования помета, так и после смены зубов. Генетическая обусловленность этих аномалий не вызывает сомнений.
Врожденные отклонения от нормального числа премоляров, их размеров и формы обнаруживаются у всех представителей отряда хищных. Можно выделить следующие механизмы их возникновения: 1) расщепление зубного зачатка; 2) слияние зубных зачатков; 3) развитие дополнительного зубного зачатка в зубной пластинке; 4) отсутствие одного или нескольких зачатков в зубной пластинке.
Часть недостатков зубной системы оказываются связанными с излишним удлиннением лицевой части черепа, как, например, у доберманов. Этот процесс часто идет преимущественно за счет верхней челюсти и сопровождается формированием укороченной или слишком мелкой нижней челюсти. Возрастание длины челюстей приводит к появлению лишних премоляров и моляров. При слишком узкой морде, резцовый ряд нижней челюсти чрезмерно изгибается вперед. Это приводит к тому, что при недостатке места закладываются только пять или даже четыре резца. Подобное явление отмечается и у собак мелких пород, имеющих относительно крупные зубы. Замечено, что у пород, имеющих голову классического строения, например у немецких овчарок, обнаруживается меньшее количество дефектов резцов.
Резцовая олигодонтия может проявляться по разному:
• зубы нормального размера, но количество их меньше — четыре или пять вместо положенных шести;
• два зуба срастаются вместе от корней до верха коронок образуя один более крупный зуб нормальной формы. Если это происходит с одной стороны, то вырастает пять зубов, а если с двух — то четыре;
• два крайних зуба срастаются своими корнями, коронки зубов оказываются разделенными своими верхушками. Образуется, так называемый сдвоенный зуб.
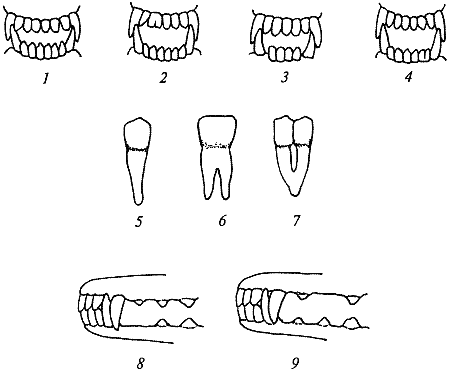
Рис. 21. Олигодонтия по резцам и премолярам: 1 — челюсти с нормальным количеством резцов; 2 — пять резцов в верхней челюсти, второй зуб слева сдвоенный, образован путем срастания коронок; 3 — челюсти с уменьшенным количеством резцов: пять резцов в верхней челюсти и четыре в нижней; 4 — пять резцов в нижней челюсти. Крайний правый зуб образован в результате срастания корней двух соседних; 5 — нормальный резец; 6 — сдвоенный резец, образованный срастанием коронок и частично корней; 7 — сдвоенный резец, образованный срастанием корней двух соседних зубов; 8 — олигодонтия по премолярам нижней челюсти. Отсутствует Р1; 9 — олигодонтия по премолярам верхней челюсти. Отсутствует Р2
Многозубость или иначе полиодонтия встречается в форме типичной, или атавистической, для которой характерно наличие избыточных зубов внутри зубного ряда и которая бывает почти физиологичной и атипичной, когда сверхкомплектные зубы вырастают вне зубных лунок, иногда даже вне ротовой полости. Псевдополиодонтия чаще всего связана с сохранением молочных зубов. Сверхкомплектные зубы могут быть представлены в виде сдвоенного зуба.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:06 | Сообщение # 99 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| 
Рис. 22. Полиодонтия: 1 — по резцам; 2 — по клыкам, образованная невыпавшим молочным клыком
Полиодонтия часто являются следствием недостаточности щитовидной железы. Некоторые породы имеют повышенную частоту полиодонтии по резцам.
Иногда у собак карликовых и той-пород зубы бывают слишком крупными по отношению к челюсти и не могут расположиться в ровную линию, что приводит к возникновению, так называемой, нерядности. Нерядность может быть связана, также, и с недоразвитием нижней челюсти. Как показывает опыт, степень наследственной обусловленности нерядности достаточно высока.
В некоторых случаях встречается, так называемый, перекос, проявляющийся в наклоне зубного ряда в определенную сторону. Большинство заводчиков и владельцев собак склонны объяснять это травмой или склонностью собаки грызть кость или игрушку одной стороной челюсти, а также регулярным потягиванием поводка или палки в игре. Однако, как показывают генетические исследования, данный дефект или предрасположенность к нему имеют определенную наследственную детерминированность.
У породных собак, особенно мелких пород, достаточно часто наблюдаются отклонения в прорезывании зубов. Например, постоянные зубы прорезываются при невыпавших молочных, и образуется двойной комплект зубов, чаще всего клыков, или даже, полный двойной ряд зубов .
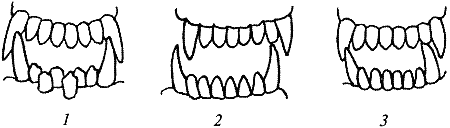
Рис. 23. Нерядность резцов и перекос челюстей: 1 — нерядность резцов нижней челюсти; 2, 3 — перекос челюстей
Подобные аномалии характерны для нарушений деятельности системы гипофиз — щитовидная — паращитовидная железа, что также встречается при несбалансированности генома. Иногда при смене молочных зубов задерживается появление премоляров, и, реже, моляров и, несмотря, на героические усилия владельца щенка, зубы все же не появляются и рентгенограмма показывает, что не были заложены даже их зачатки. Нельзя говорить о моногенном или полигенном наследовании этих нарушений, скорее этот факт говорит о том, что зубная система в процессе формирования чрезвычайно чувствительна как к повреждающим моментам, так и к несогласованной деятельности различных частей генома. В подтверждение теории несбалансированности говорит тот факт, что часто нарушения зубной системы возникают при скрещиваниях собак определенных линий.
При детальном исследовании зубной системы иногда обнаруживается ненормальное сближение корней зубов — конвергенция корней . При этой аномалии не происходит рассеяния давления производимого корнями зубов на ткани челюсти, а наоборот, возникает сильное локальное давление, со временем приводящее к трофическим нарушениям в деснах и окружающих тканях. Дивергенция корней, или иначе расхождение коронок, возникает в период смены молочных зубов в результате задержки их выпадения и приводит к развитию редкозубости. Довольно часто встречается ретенция зуба — при этом зуб находится не в зубном ряду, а в толще челюстной кости.
Таким образом, форма прикуса и всякие отклонения от полной зубной формулы в большинстве случаев определяются полигенной системой, что может быть рассмотрено как показатель неудачной комбинации блоков генов в генотипе, способной повлечь за собой множественные нежелательные последствия. Носителей этих аномалий желательно исключать из разведения.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:07 | Сообщение # 100 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Форма ушей
Длительная селекционная работа, проводимая на собаках, привела к формированию множества форм ушей. Можно выделить следующие три группы ушей: стоячие, висячие и полустоячие.
Стоячие уши направлены концами вперед и вверх. Соразмерно голове собаки стоячие уши могут быть большими или малыми. Уши могут иметь более или менее заостренные концы, быть близкими по форме к равнобедренному треугольнику с основанием короче боковых сторон, или напоминать равносторонний треугольник. Концы стоячих ушей могут быть направлены в разные стороны, образуя развешенные или сближенные уши.
Полустоячие уши имеют крепкие хрящи только в нижней половине уха, вторая же половина, в силу мягкости хряща, опускается вниз или в сторону.
Висячие уши бывают двух видов: висячие на хряще с крепким у основания хрящом, поддерживающем ухо на линии лба (ухо эрдель-терьеров) и висячие — хрящи которых мягки и уши, в силу своей тяжести, свисают по обеим сторонам головы собаки (у различных пород гончих и легавых). Висячие уши также могут быть длинными и короткими, а также узкими и широкими с закругленными или острыми концами.
Независимо от постава, уши могут быть высоко и низко посаженными.
Велико разнообразие и в степени оброслости ушных раковин шерстью. Стоячие уши обычно покрыты короткой прямой шерстью; висячие могут быть покрыты как короткой, так и длинной шерстью.
Наследование формы и размера ушной раковины полигенно, но можно выделить определенные локусы, оказывающие значительное влияние на отдельные детали ее строения.
Так показано, что при скрещивании собак с висячими ушами с собаками со стоячими ушами, наблюдается неполное доминирование со стороны висячего уха.
HH — висячее ухо; hh — стоячее ухо; Hh — полустоячее ухо.
В этом же локусе присутствует и аллель доминирующий полностью над H и h — Hа — полустоячее ухо типа «колли». При этом собаки с генотипами HаHа, HаH и Hаh фенотипически неразличимы. Таким образом, в популяциях колли, терьеров и других пород, могут присутствовать все три аллеля, что приводит к периодическому выщеплению собак со стоячими ушами.
Очевидно, такой тип наследования характерен для внутрипородных популяций. В случае, если ушные раковины скрещиваемых пород сильно различаются по размеру, эта закономерность может быть утеряна.
Также показано неполное доминирование и взаимозависимость размеров ушей. Если висячее ухо маленькое, то доминирует стоячее ухо, но получившееся стоячее ухо больше, чем у исходной формы. Если висячее ухо большое или длинное, то стоячее ухо доминирует не полностью. Но, тем не менее, гибриды F1, полученные от скрещивания пуделей с волками и шакалами («пуво» и «пуша»), имели стоячие уши. Гибриды F2имели уши разнообразной формы и размера.
Авторы наблюдали целый ряд межпородных гибридов и проследили наследование нескольких признаков. Гибриды F1пуделя и английского бульдога (с ушами розовидной формы) имели уши по форме и размеру похожие на пуделиные, однако более высоко посаженные. Гибрид F1пуделя и фокстерьера имел полустоячие уши в типе керриблютерьера. Гибриды F1карело-финской лайки и фокстерьера имели стоячие уши. Гибриды F1эрдельтерьера и ризеншнауцера имели уши по типу близкие к ушам ризеншнауцера.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:08 | Сообщение # 101 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Окраска глаз собаки
Окраску глаз собак можно условно разделить на две категории: нормальную и аномальную.
Нормальная окраска глаз собаки зависит от локусов окраски шерсти Е,В,D и может быть более или менее темной — темно-карей или черной, ореховой или карей и желтой. Эти окраски соответствуют трем аллелям Jr; Jrm и iry, которые, по-видимому, не полностью доминируют одна над другой. У собак с ослабленной пигментацией, гомозиготных по рецессивному аллелю dd — цвет глаз дымчатый, что мы иногда наблюдаем у серорожденных пуделей и догов. У рецессивных гомозигот bb (коричневых) при генотипе Jr — цвет глаз каштановый, при irm — светло-ореховый и iry — светло-желтый.
При В — Jr — дает почти черный цвет глаз, irm — темно карий, iry — карий.
К аномальным окраскам глаз можно отнести рубиновоглазие, гетерохромию радужной оболочки и альбиностические. Подобные окраски глаз не представляют собой патологии, и, чаще всего, не оказывают влияния на остроту зрения собак.
Рубиновоглазие, проявляется в том, что при освещении под определенным углом в глазах собаки появляется рубиново-красный отсвет. Этот феномен не зависит от цвета глаз. Исследования показали, что у таких собак наблюдается своеобразное расположение пигмента в радужной оболочке глаза, которое не влияет на зрение собаки и передается как рецессивный признак. Аллель определяющий его развитие обозначен как р. Отмечены случаи сочетания рубиновоглазия со светло-серым окрасом шерсти у пекинесов.
У альбиносов полностью отсутствует пигмент в радужной оболочке глаза и, в связи с тем, что через ее толщину просвечивают сосуды она имеет красный цвет. Альбинизм глаз различной степени отмечался среди фокстерьеров, мальтийских болонок, боксеров, догов и далматинов. Но в целом альбиносы среди собак весьма редки.
Гетерохромия радужной оболочки («беловзор», «сорочий глаз») связана с аномалией радужной оболочки, которая у животных состоит из трех слоев различной толщины: переднего и заднего и лежащей между ними стромы. В нормальном глазу строма более или менее пигментирована за счет насыщенности пигментными клетками, также за счет пигментного эпителия заднего слоя радужной оболочки, что и определяет естественный цвет глаз. Этот самый задний слой у «сорочьего глаза», в отличие от альбиностического нормально пигментирован. Лежащие впереди его пигментные клетки стромы имеют веретенообразную форму, они сильно разветвлены, заполнены пигментом и снаружи уплотняются, образуя густую сеть — пограничный слой. Лишь недостаток пигмента в строме радужной оболочки ведет к клиническому проявлению гетерохромии радужной оболочки, называемой «беловзор», или «сорочий глаз». Полное отсутствие пигментных клеток стромы обуславливает полную и частичную гетерохромию радужки. Из-за большой рыхлости соединительной ткани глаза радужная оболочка «сорочьего глаза» выглядит голубоватой, в связи с этим точнее его было бы назвать «галочьим». Как при полной, так и неполной гетерохромии радужки, участки, лишенные пигмента, обычно отличаются фарфоровой белизной с серо-голубой зоной зрачка и краем склеры; но и здесь при сужении зрачка серо-голубой оттенок усиливается.
В зависимости от локализации и формы различают:
1) одностороннюю частичную гетерохромию радужки (правостороннюю или левостороннюю);
2) одностороннюю полную гетерохромию радужки (правостороннюю или левостороннюю);
3) двустороннюю частичную гетерохромию радужки;
4) левостороннюю полную гетерохромию радужки с правосторонней частичной или наоборот;
5) двустороннюю полную гетерохромию радужки;
Почти половина всех случаев связана с редукцией пигмента в вентральной части глаза, примерно, 30%, — в медиальной, и лишь относительно редко встречаются участки дорсальной, латеральной или циркулярной локализации.
Как показало большинство исследований, представляется, что гетерохромия не оказывает никакого влияния на остроту зрения и светочувствительность. Такой глаз во всех случаях реагирует на свет непосредственно обычным резким сужением зрачка. В сумерках зрачок выглядит совершенно нормально. У собак описаны случаи, когда с гетерохромией радужки нередко сочетались другие тяжелые нарушения глаз, как микрофтальмия и колобома, а также глухота.
У различных пород собак гетерохромия радужки сочетается с пятнистым или мраморным окрасом. Однако, между этими признаками не существует абсолютной корреляции. Гетерохромия наследуется обычно как простой аутосомно-рецессивный признак, однако некоторые авторы описывают его как аутосомно-доминантный.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:09 | Сообщение # 102 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Шерстный покров
В процессе породообразования шерстный покров собак претерпел значительные изменения. Среди них можно встретить как шерсть близкую к «дикому» типу, разные варианты мягкой, жесткой и полужесткой, длинной и совсем короткой и, даже, практически голых.
Волосы собак могут иметь разнообразную форму. Прямой волос имеет прямой, лишь с небольшим изгибом в месте расширения стержень; изогнутый — с постепенным изгибом в одну сторону: волос с надломом — с резким переломом в одну сторону ; волнистый — имеет стержень, отклоняющийся от прямой оси в обе стороны; кольцеобразный или спиральный — закручен в одну сторону и образует плотные кольца или спирали.
Часть пород сохраняет «дикий» или скорее «нормальный» тип шерсти: двухъярусный мех и неодинаковую длину и фактуру шерсти на разных частях тела.
У собак и диких собачьих можно выделить, по крайней мере, три разновидности шерсти дикого типа:
1) «Нормальный» — шерсть средней длины. Присутствует у большинства видов средней полосы — волков, шакалов, лесных и дальневосточных кошек и у многих пород естественного облика: лаек, овчарок, гончих средней полосы и т. д.
2) «Укороченная шерсть дикого типа» — у динго и новогвинейской собаки и оцелота среди диких форм. У собак укороченную шерсть дикого типа имеют мопсы, стаффордширские терьеры, бульдоги и т. д.
3) «Удлиненная шерсть дикого типа» — встречается в природе у лисиц, песцов, енотовидных собак, красных волков. Среди домашних собак у самоедов, вольфшпицев, кесхаундов, чау-чау, колли, кавказских овчарок и т. п.
Всю массу короткошерстных собак можно разделить на собак с укороченной шерстью дикого типа с выраженным подшерстком и волосами всех типов и гладкошерстных собак. Гладкошерстные собаки обычно не имеют подшерстка, при этом покровные волосы в ряде случаев имеют незначительные отличия на разных частях тела. У некоторых собак эти различия практически нивелируются.
Очевидно, гладкошерстность может быть определена иными аллелями, чем короткошерстность дикого типа. Короткошерстность дикого типа является доминантной по отношению к длинной шерсти, а гладкошерстность — рецессивной.
Исследователями описан ряд аллелей, определяющих некоторые свойства шерстного покрова собак. Однако необходимо отметить, что их обозначения и трактовка разными авторами не всегда совпадают.
Длина шерсти обозначается символом L.
L — шерсть «дикого» типа; l 1— длинная шелковистая шерсть; l2 — гладкая короткая шерсть. По-видимому, доминирование в этом локусе имеет следующий порядок: L› l 1 › l2.
Аллель l1 продлевает период роста волоса, что и способствует увеличению его длины. Возможно, что вариации длины шерсти и особенностей строения определяются рядом генов-модификаторов данного аллеля.
Длинношерстные породы могут сохранять выраженные отличия покровных волос и подшерстка (колли, ретриверы, ньюфаундленды и т. д.) или почти не иметь подшерстка как, например, сеттеры. У некоторых из них остевые волосы стали настолько тонкими, что практически не отличаются от подшерстка (йоркширские терьеры, афганы). У собак типа пуделей, командоров и пули остевые волосы и подшерсток имеют одинаковую длину и перевиваются друг с другом.
Распределение волос разной длины по корпусу у длинношерстных собак также имеет разный характер. Некоторые из них сохраняют дикий тип оброслости с длинной шерстью на теле, хвосте и на бедрах и укороченной на морде и ногах (колли, шпицы). Другие имеют равномерную оброслость по корпусу, ногам и морде (комондоры, пудели, мягкошерстные терьеры, ши тцу), так называемую брудастость. При скрещивании собак этих двух типов доминирующим оказывается второй тип.
Очевидно, можно предположить наличие особого локуса, ответственного за характер оброслости. Мы предлагаем обозначить данный локус как W (wild) и выделить, по меньшей мере, два аллеля W — равномерную оброслость по всему телу собаки и w — обусловливающий дикий тип оброслости.
Курчавая шерсть может быть также очень разной: волосы могут закручиваться в спираль, или кольца, быть похожими на металлическую стружку или равномерно волнистыми. Возможно, что курчавая шерсть ирландских водяных спаниелей и ретриверов, а также волнистая шерсть кокеров и других пород может определяться разными генами.
Скрещивание пуделей с кокерами и другими породами с подобной шерстью дает в первом поколении собак с шерстью типа кокера. Это показывает, что данные аллеи относятся в одному локусу и волнистая шерсть доминирует над скрученной.
Таким образом, можно унифицировать обозначения данных аллелей следующим образом: wa — неволнистая шерсть, wa1 — волнистая шерсть , wa2 — скрученная (спиральная) шерсть. Доминирование аллелей в этом локусе очевидно носит следующий характер: wa›wa1› wa2.
Универсальным для всех жесткошерстных пород собак является ген жесткошерстности Wh. Рецессивный аллель этого локуса обусловливает развитие шерсти дикого типа. При генотипе llWh — волосы длиннее, чем при LLWh-, но короче, чем при llwhwh. Порядок доминирования аллелей в этом локусе: Wh›wh.
Шерсть такого типа встречается у собак с жесткой прямой шерстью — шнауцеров, брюссельского гриффона, некоторых терьеров, жесткошерстной таксы, дратхаара. Метисы собак этих пород большей частью также имеют шерсть данного типа. Многие курчавошерстные терьеры при скрещивании с прямошерстными породами полностью утрачивают завиток и их метисы выглядят так же, как и метисы собак с жесткой прямой шерстью.
Жесткие волосы часто бывают извитыми или курчавыми, поэтому иногда бывает трудно провести четкую грань между жесткошерстными, полужесткошерстными и курчавыми собаками.
Курчавая, скрученная и волнистая шерсть в сочетании с аллелями Wh и wh и L и l, определяют шерстный покров целого ряда пород. Пудели, очевидно, имеют генотип Wh-llkk или WhWhllkk. При скрещивании их с волками или собаками с диким типом шерсти получаются собаки с довольно длинной прямой и достаточно жесткой шерстью, скрученность, курчавость или волнистость шерсти полностью исчезает. Метисы различных спаниелей, полученные при скрещивании с длинношерстными собаками, имеют весьма характерный фенотип и шерсть близкую по типу к спаниелям. Метисы спаниелей и короткошерстных собак имеют обычно шерсть дикого типа.
Мы предлагаем следующую систему формирования шерстного покрова собак (табл. 10).
Кроме вышеописанных аллелей, обусловливающих развитие тех или иных характеристик шерстного покрова существует масса факторов, как наследственного, так и ненаследственного характера, также оказывающих влияние на формирование типа шерсти у животного.
К наследственным можно отнести множество генов-модификаторов, влияющих на развитие и соотношение волос разного типа (подшерстка и остевых волос), а также возможную неполную пенетрантность и экспрессивность описанных аллелей, вкупе обеспечивающих большую широту нормы реакции, что, в свою очередь, способствует многообразию проявлений модификационной изменчивости шерстного покрова.
К ненаследственным можно отнести влияние климатических условий, тип кормления, наличие или отсутствие стрессорных факторов и т. д. Даже внутри одной породы могут встречаться собаки с разной шерстью. Многое зависит и от способов ухода за ней. Так, например, у одного и того же эрдельтерьера после стрижки или щипки сформируется совершенно разный волосяной покров, отличающийся и по структуре и, даже, по интенсивности окраски.
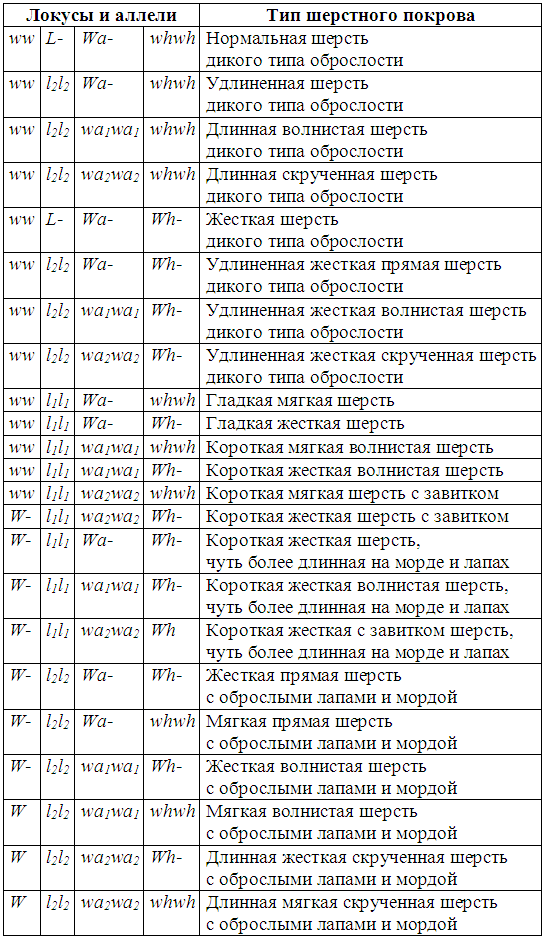
Таблица 10. Система формирования шерстного покрова собак
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:09 | Сообщение # 103 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| У собак существует целый ряд мутаций, приводящих к развитию безволосости и отмеченных в разных точках земного шара. Встречаются разные обозначения безволосости собак. Так Н.А. Ильин (1932) обозначает эти аллели следующим образом: N — безволосость; n — нормальное состояние шерстного покрова. Робинсон обозначает ген безволосости как Hr , и hr как нормальное состояние шерстного покрова. Аллели безволосости в гомозиготном состоянии приводит к внутриутробной гибели его носителей. Все живущие голые собаки являются гетерозиготными и при скрещивании между собой дают в каждом помете гомозиготных особей, имеющих нормальный шерстный покров, так называемых «пуховок».
У мексиканских и китайских голых собак редкий шерстный покров есть только на голове и ногах, на теле присутствуют лишь отдельные волосы, возможно типа вибриссов. Кожа голых собак испещрена рисунком мягких складок, увеличивающих истинную поверхность кожи и выполняющих множественные защитные функции.
Сама по себе их кожа достаточно тонкая, слой подкожной жировой клетчатки также невелик. Отсутствие волос и сравнительно тонкая кожа, очевидно не могут обеспечить достаточно хорошую термоизоляцию, поэтому голые собаки легко мерзнут и нуждаются в предохранении от охлаждения.
Безусловно аллели безволосости, как и аллели дикого типа, обладают плейотропным действием. Сам факт внутриутробной гибели щенков, гомозиготных по гену безволосости, говорит о глубоком нарушении органогенеза. Плейотропное действие гена безволосости проявляется и в формировании определенного породного фенотипа. В частности зубы у голых собак по своему строению во многом отличаются от зубов собак других пород.
Очевидно, бесшерстность китайских и мексиканских голых собак обусловлена разными аллелями. Экспериментальные вязки мексиканских голых собак с беспородными собаками, подходящего размера и фенотипа (за исключением бесшерстности) приводят к рождению в пометах голых щенков по фенотипу не отличимых от породных, в то же время, среди пуховок наблюдается огромный разброс по фенотипу. Пуховки мексиканских голых собак полученные в породных пометах также весьма разнообразны и не допускаются к выставкам и племенному разведению.
Китайские голые собаки генетически более стабильны и их пуховки имеют определенный фенотип, что позволяет им участвовать в выставках и племенной деятельности.
Кроме доминантного гена безволосости с плейотропным действием у собак встречается и рецессивный ген безволосости, обусловливающий развитие шерстного покрова у американских голых терьеров. Щенки американского голого терьера рождаются опушенными и теряют шерсть к 6–8 неделям жизни. Интересно отметить, что собаки этой породы не имеют аномалий зубной системы, типичных для других голых собак.
Очень интересная особенность шерстного покрова встречается у риджбеков. У собак этой породы на спине имеется гребень, образованный волосами, растущими в направлении, противоположном росту остальных волос, так называемый «ридж». Иногда гребней бывает два. Стандарт породы требует, чтобы гребни были четко обозначены, симметричны и сужались к хвосту. Этот признак наследуется рецессивно и обозначается символом ds.
Как показывают исследования, проведенные на разных видах животных, оказывается довольно эффективной массовая селекция по длине или густоте шерсти. Однако между ними существует отрицательная корреляция, так как высокая частота расположения волосяных фолликулов уменьшает площадь их питания, что, и регламентирует рост шерсти в длину. Подобную корреляцию можно отметить и у собак — длинная шерсть йоркширских терьеров, ши тцу, два типа шерстного покрова встречается и у пуделей: густой с хорошо выраженным подшерстком и медленно растущий, не достигающий значительной длины и более редкий, но отличающийся большей длиной.
Рост шерсти также зависит от физиологического состояния организма, уровня кормления и значительно замедляется во время беременности и лактации.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:10 | Сообщение # 104 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Генетика поведения собак
Несмотря на все многообразие и сложность поведения собаки, его наследование подчиняется тем же закономерностям, что и морфологические признаки. Разнообразие пород, отличающихся друг от друга формами поведения, издавна привлекало внимание многих ученых к изучению генетики поведения. Существование разных пород собак представляет собой рекордный по длительности селекционный эксперимент, демонстрирующий как никакой другой, фантастический размах внутривидовой изменчивости и по морфологическим признакам, и по признакам поведения.
Изучению генетики поведения собаки посвящены работы таких всемирно известных исследователей, как Скотт, Фуллер, Уитни, Мархлевски, Крушинский и многих других ученых.
Хорошо изучено наследование оборонительных реакций, много научных работ посвящено генетике наследования охотничьего поведения. Показана генетическая обусловленность способности к апортировке, плаванию, лаю.
Несмотря на свою кажущуюся сложность поведения, каждый его акт может быть разделен на отдельные составляющие, наследование которых осуществляется по моногенному типу. Как показали исследования проведенных на собаках пород бассенджи и английский кокер, отдельные компоненты комплекса оборонительного поведения щенков такие как, стремление убежать, визг, мочеиспускание от страха, переворачивание на спину и т. п., детерминированы всего лишь одной или двумя парами генов.
Можно выделить следующие основные направления изучения наследования закономерностей поведения:
1. Наследование специфических свойств поведения, например, злобности, пугливости, некоторых особенностей поведения охотничьих собак, апортировочной реакции и т. п.
2. Наследование общих свойств нервной системы, например, возбудимости или поведения в зависимости от пола.
3. Исследование различий в социальном поведении собак разных пород.
4. Исследование корреляций между физическими и поведенческими чертами.
5. Комплексный анализ поведения разных пород собак с целью выбора наиболее приемлемой породы для определенных целей с применением компьютера. Кластерный анализ.
6. Изучение поведения гибридов собак с дикими животными: волками, шакалами и койотами.
Поведение играет исключительно важную роль в эволюции и зависит от естественного отбора. Изучение элементарной рассудочной деятельности диких и доместицированных форм одного вида показало, что одомашненные животные решали предлагавшиеся им задачи несколько хуже, чем дикие. Это вполне понятно, так как в природе выживают, прежде всего, те особи, которые способны находить правильный выход их экстренно сложившихся ситуаций. Очевидно, разум является одним из факторов, способствующих лучшему выживанию в природных условиях. В процессе доместикации наиболее умные животные зачастую напрямую выбраковываются человеком или убегают на свободу. Они оказываются наиболее предприимчивыми и самостоятельными и поэтому менее удобными в содержании в неволе, что часто приводит к созданию неожиданных ситуаций.
При содержании шоу-собак в питомниках идет отбор на тех, которые спокойнее всего реагируют на содержание в вольерах или клетках, позволяют проводить с собой разнообразные манипуляции, например причесывание, тримминг и стрижку, способны не отвлекаясь выдерживать многочасовое стояние на столе в процессе подготовки к рингу и т. п. Можно предположить, что такая селекция идет не в пользу наиболее умных и любознательных собак.
Содержание в домашних условиях способствует отбору аккуратных животных с выраженным гигиеническим поведением. В питомниках зачастую на этот фактор не обращают должного внимания, в результате чего многие владельцы собак, питомникового разведения, а также владельцы их потомков жалуются на то, что их оказывается трудно, а порою и невозможно, приучить к поддержанию чистоты в квартире. При этом более жесткому отбору подвергаются крупные собаки, вследствие чего данный недостаток встречается среди них реже, чем среди мелких.
Серьезную генетико-поведенческую проблему представляет спонтанная агрессия некоторых собак, возникающая на фоне периодических выбросов адреналина. Такая собака при малейшем возбуждении, возникшем, например, во время игры или прихода гостей, внезапно начинает кусать окружающих и, в первую очередь, своих хозяев. Этот признак имеет четкую наследственную обусловленность и широко распространен среди собак некоторых пород. Так, например, эта немотивированная агрессия вызывает большую озабоченность у разведенцев кокеров во всем мире.
Очень часто разнообразные аномалии поведения, как то, повышенная нервозность, аномальная ласковость, клаустрофобия и др., служат маркерами нестабильности генома и являются признаками гибридного дисгенеза.
Поэтому отбор по поведению должен занимать одно из первых мест в селекционной работе и особи с его нарушениями должны самым жестким образом выбраковываться из числа производителей, как бы ни был великолепен их экстерьер. В погоне за улучшением внешнего облика необходимо помнить, что основное назначение собаки — быть умным и приятным в общении другом.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
| Svarta | Дата: Четверг, 26.04.2012, 19:12 | Сообщение # 105 |
|
Бессмертный
Группа: Администраторы
Сообщений: 7824
Статус: Offline
| Генетические аномалии и пороки развития
На основании анализа наследственно обусловленных аномалий их можно разделить на две группы.
Генетические аномалии представляют собой наследственно обусловленные, с точки зрения здоровья популяции и племенного использования, отклонения от типичного (от нормы), в возникновении которых определенную роль сыграл генотип животного. Большая часть генетических аномалий существует в популяциях в виде рецессивных летальных или сублетальных генов, находящихся в гетерозиготном состоянии. Подобных генов у собак описано меньше, чем у других животных, однако это говорит лишь о меньшей их изученности.
Породные аномалии представляют собой отклонения от существующего породного типа, часто возникающие вследствие некорректного использования различных методов разведения.
Генетические аномалии — это морфофункциональные нарушения в организме животного, возникающие вследствие генных и хромосомных мутаций. Генные мутации могут нарушать морфогенез органов и тканей на разных этапах, отсюда столь широкий спектр врожденных аномалий, связанных с изменениями молекулы ДНК. Изменения числа хромосом в клетках или их структуры обычно приводят к прекращению развития эмбриона или рождению особей с тяжелыми пороками развития, нарушению у животных воспроизводительных функций. Однако, далеко не все генетические аномалии представляют собой патологию. Часто под «аномалией» понимается отклонение от нормального фенотипа. Например, иной цвет глаз, появление нетипичных для породы пятен, отклонения в количестве зубов или непредусмотренная стандартом фактура и длина шерсти.
Большая роль в этиологии врожденных аномалий принадлежит летальным и сублетальным генам. Так, у человека ими обусловлено свыше 2000 аномалий. Много таких признаков описано и у домашних животных разных систематических групп.
Развитие многих аномалий определяется взаимодействием факторов окружающей среды и генотипа. Это, так называемые, наследственно-средовые аномалии, которые, по-видимому, контролируются полигенной системой. Фенотипическое проявление этих признаков зависит от количества мутантных генов, обуславливающих аномалию. Существует понятие порога действия таких генов, что соответствует их числу, то есть силе кумулятивного действия, необходимого для проявления аномалии.
Если число мутантных генов ниже порогового, животное останется фенотипически нормальным. В то же время высота порога действия зависит от условий среды.
В некоторых случаях фенотипически сходные аномалии имеют разную генетическую детерминированность. В других случаях возникновение фенотипически сходных аномалий у особей с определенными генотипами может происходить под действием внешней среды. Такие аномалии называются фенокопиями.
Аномалии могут возникать под воздействием повреждающих факторов внешней среды — тератогенов. Часто они оказываются сходными с генетически обусловленными дефектами. Так например, «волчья пасть» может возникнуть как при тератогенном воздействии, так и быть наследственно обусловленной.
Список основных генетических аномалий собак
(Подробнее об этом можно прочесть в книгах: Московкиной Н.Н. и Сотской М.Н. «Генетика и наследственные болезни собак и кошек» М., 2000, а также М.Б. Уиллис «Генетика собаки», М., 2000).
Аномалии внешних покровов: пузырчатка (пемфигус), красная волчанка, токсический эпидермальный некролиз, полиморфная эритема, холодовой агглютинит, кожный васкулит, узловатый панникулит, опухоли — липомы, папилломы и бородавки, злокачественные новообразования, кожная астения (дерматоспараксия), ихтиоз, зависимый от витамина А дерматоз, врожденный гипотрихоз, Х-алопеция собак северных пород, рецидивирующая алопеция боковой поверхности тела собак, алопеция цветных собак (синдром голубого добермана), черный акантоз, цинковый дерматит, солнечный дерматит мочки носа (нос колли), акродерматит, дерматит вельштерьеров, юношеский целлюлит, семейный дерматомиозит, дермоидные кисты.
Аномалии развития нервной системы и органов чувств: дисплазия центральной нервной системы, гидроцефалия, микроцефалия, эпилепсия, рефлекторная эпилепсия, нарколепсия, мозговая грыжа, менингоэнцефалоцеле, спинальная атаксия, миотония, судороги скотч-терьеров, тремор белых собак, мышечная дистрофия лабрадоров (миопатия), мышечная дистрофия, синдром атрофии мышц спины, миастения, дегенеративная миелопатия, ганглиозидоз GM1, ганглиозидоз GM2, сфингомиелоноз, полинейропатия, сенсорная невропатия, прогрессирующая аксонопатия, гигантская аксональная невропатия, синдром пошатывания, гипомиелиногенез, хроническая дегенеративная радикуломиелопатия, мутация демиелинизации, болезнь Краббе, укорочение позвоночника, дисплазия спинного мозга и врожденное расщепление позвоночника, синдром неустойчивого позвонка — шейная спондилопатия, квадриплегия и амблиопия.
Аномалии уха: аномалии наружного и внутреннего уха, глухота далматинов, поражение вестибулярного аппарата.
Аномалии глаз: микрофтальмия, анофтальмия (синдром непрорезывания глаз), аномалии глаз колли, глазные дермоиды и дермоидные кисты, колобома, эпибульбарная дерматофиброма, врожденный заворот век (энтропия), эдема роговицы, дистрофия роговицы, мумификация роговицы, перестирующая перепонка зрачка, катаракты, афаксия (афакия), (эктопия), вывих или подвывих хрусталика, первичная глаукома, вторичная глаукома, прогрессирующая атрофия сетчатки — радиальная, центральная, отслойка сетчатки, дисплазия сетчатки, расстройство сумеречного зрения, аномалии зрительных путей.
Аномалии опорно-двигательного аппарата: волчья пасть (палатосхиз, расщелина неба), расщепление верхней губы, верхней челюсти и мочки носа, «двойная мочка носа», прогения, прогнатия, остеопатия нижней челюсти, краниомандибулярная остеопатия, расщелины черепа, синдром конского хвоста, деформирующий спондилез, смещение межпозвонковых дисков, таксиный паралич, остеохондроз, отсутствие хвоста, короткий изогнутый хвост, полупозвонок, клиновидный позвонок, абрахия, биторакальная эктромелия, микромелия, синдактилия, брахидактилия, полидактилия, некроз пальцев лап, подвывих запястья, вывих коленной чашечки, множественная фиброзная дистрофия, дисплазия тазобедренного сустава, болезнь Лагга-Кальва-Пертеса, некроз головки бедренной кости, искривление локтевой кости, дисплазия локтевого сустава, дисплазия локтевого бугра, изолированный локтевой отросток, изолированный медиальный надмыщелок, подвывих локтя, вывих локтя, смещение головки лучевой кости, полиартродисплазия, артрогриппоз, паностеит.
Заболевания крови: гемофилия А, гемофилия В, неостаточность фактора свертываемости крови XII, недостаточность фактора VII, болезнь Стюарта-Прауэра, болезнь Виллебранда (ангиогемофилия), циклическая нейтропения (болезнь серых колли), полицетемия, лейкозы, тромбоцитоз, аномалия Пельгера-Хьюэта, синдром Чедиак-Хигаши, острая интермиттирующая порфирия, эритропоэтическая порфирия.
Наследственные заболевания иммунной системы: тиреоидит, системная красная волчанка, ревматоидный артрит, инфаркт миокарда, миастения, некоторые формы нефритов, аутоиммунная гемолитическая и пернициозная анемии, идиопатическая тромбоцитопения, пузырчатка (пемфигус), комбинированный иммунодефицит (СID), ангионевротический отек и крапивница, аллергии.
Опухоли: лейкозы, миеломы, нейрофиброматоз, липоматоз (множественные липомы), полипоз желудочно-кишечного тракта, и ретинобластома, пигментная ксеродерма, аденома эндокринных желез, рак толстой или прямой кишки, желудка и двенадцатиперстной кишки, рак молочных желез и матки, рак предстательной железы, щитовидной железы, опухоли почек, яичников, семенников (семинома), гортани и т. д.
Аномалии сердечно-сосудистой системы собак: незаращение боталлова протока, тетрада Фалло, стеноз (сужение) легочной артерии, правосторонняя дуга аорты, двойная дуга аорты, дивертикул пищевода, сужение (стеноз) аорты, недостаточность атриовентрикулярных клапанов, идиопатический эндокардиальный фиброэластоз, кардиомиопатии.
Аномалии пищеварительной системы: язвенный стоматит, неправильное стирание зубов, парадонтоз, гиперпластический гингивит, ахалазия пищевода, дисфункцию пищевода, дивертикул пищевода, хронический склерозирующий панкреатит (панкреоцирроз), наследственная атрофия поджелудочной железы, инсулинома, экзокринная недостаточность поджелудочной железы, хронический склерозирующий панкреатит, кистозный фиброз поджелудочной железы, мальабсорбция, перианальная фистула, атрезия анального отверстия, внешний кишечник.
Аномалии выделительной системы: аномалии количества почек, аномалии положения почек, гипоплазия почек, нефропатия, почечный амилоидоз, поликистоз почек, семейно обусловленный нефроз, наследственная цистинурия, мочекаменная болезнь.
Аномалии обмена веществ: врожденный медный токсикоз, нарушения обмена цинка, глютеновые энтеропатии, фукозидоз, гликогенная болезнь, ганглиозидоз, недостаток энзима каталазы, недостаток фермента пируваткиназы, нарушение процесса усвоения витамина В12, мукополисахаридоз, сахарный диабет, истинная карликовость, хондродистрофия, ложная непропорциональная карликовость, ложная пропорциональная карликовость.
Аномалии пигментации: альбинизм, болезнь серых колли, пигментные пятна.
Аномалии половой системы: пороки развития половых желез, истинный гермафродитизм, ложный гермафродитизм, синдром тестикулярной феминизации, крипторхизм.
Аномалии развития плода: мумификация плода, врожденная водянка плода и плодных оболочек, врожденная контрактура мышц плода.
Грыжи: паховая, мошоночная, промежностная и диафрагмальная грыжи.
Друг это тот , кто все про тебя знает , и все равно любит .
ВСЕ ЧТО ГОВОРЯТ ОБО МНЕ ЗА МОЕЙ СПИНОЙ, СЛУШАЕТ ТОЛЬКО МОЯ ЖОПА.......!!!!!!! Ф. Раневская
|
| |
|
|
|









